
Introduction
Wheat grain is a staple food for almost 2.5 billion people in 89 countries due to its adequate nutritional value, storage capacity, and processing capabilities (CGIAR 2022).1 The pivotal challenge is to enhance the production and productivity of the crop, particularly in the areas under climatic vagaries. According to the Intergovernmental Panel on Climate Change’s fifth assessment report, the average global surface temperature of the land and ocean has risen by 0.85°C during the past three decades.2 By the end of the 21stcentury, the increase in average global temperature by 1–40C is predicted.3 The reproductive stages of wheat are more susceptible to high temperatures than their vegetative counterparts, like many other cereals. The primary cause of the abrupt yield reduction in such circumstances is pollen infertility.4-5 Due to different physiological injuries, including leaf abscission, senescence, scorching of leaves and stems, inhibition of root and shoot growth, grain damage, decreased starch accumulation, altered starch-lipid content in grains, decreased seed germination, and decrease in vigour, plant productivity is consequently reduced.6 Diminished plant productivity and shortened life cycle can be caused due to heat induced changes in respiration and photosynthesis.7 Yield loss even up to 40 % under severe heat stress conditions has been reported.8 Because of changed climatic scenario the essentially required cool period at critical growth stages for wheat crop is shortened in India and aggravates the threat of terminal heat stress.9When mature wheat crops combine with terminal heat stress (THS), the yield and quality of grains are likely to be alarmingly threatened in tropical and subtropical countries.10 The development of high yielding cultivars with climate resilience needs extensive programmes of plant breeding. It has become very common in this field to use direct selection for yield stability depended on several site trials under unfavourable weather conditions. Selection for heat tolerance genotype based on early flowering type, high pollen shedding, high spikelet fertility, low grain weight, heat susceptibility index has been rewarding.11 But the approach is time consuming and labour-intensive. As a result of poor heritability and a strong influence of several environmental factors, classical plant breeding became more challenging for abiotic stress-related traits.12 In a traditional breeding program, the selection is based on genes with significant effects that determine a distinct phenotype. Using a large population could be helpful to attain a decent likelihood of combining desired alleles at several loci in a single line for additive gene action. The lack of in-depth knowledge about the underlying molecular mechanism and appropriate techniques to address them are also the bottlenecks for crop improvement through classical breeding under abiotic or biotic stress.13 Strategic marker-assisted breeding is a successful alternative strategy that can hasten the emergence of tolerable cultivars but in-depth understanding of the genomic locus influencing the traits and the availability of relevant molecular markers is much needed.14 With the advent of the molecular marker and Quantitative Trait Locus (QTL) mapping, new breeding techniques such as marker-assisted selection and molecular breedingis becoming popular day by day.15-16 But QTL mapping alone is inefficient to handle plant’s intrinsic complexities like polygenic control, epistasis, genotype x environment (G x E) interactions etc. Some common limitations including reliability and accuracy of QTL mapping, the insufficient linkage between the marker and desired gene and the limited number of markers as well as polymorphism effects of genetic background and environmental effects on polygenes can compromise the Marker Assisted Selection (MAS) to some extent. The allohexaploid genome sequence of wheat and its genomic complexity has made the investigation of gene function more challenging.
The multidisciplinary “omics” discipline and other fields of applied biology research, uses high-throughput techniques.17 These omics have become highly popular among scientists for molecular characterization of different stress-tolerant genotypes. A good number of research papers have been published during the last decades. We intend to identify the advancements and difficulties faced by various researchers in attempts to improve wheat under terminal heat stressed condition specifically in this review. It also provided a brief summary of recent research updates on wheat omics.
Omics in plant biology research
Omics is a basket of diverse technologies that can fulfill the understanding of changes in the organism due to genetic, nutritional and environmental alteration. It provides insight into the alteration of the plant metabolism, in particular, in response to change in the micro- and macro- environment. Genomics era started with the invention of automated sequencing technologies that led to the complete genome sequencing of Arabidopsis, Rice, Soybean, Maize and many more. The physiology and adaptation of plants to any stress have been better understood with the developments in next-generation sequencing technologies, genome-scale molecular analysis, transcriptomics, and proteome profiling in the post-genomic age. Gene discovery by forward genetics is the most popular application of recently developed genomic technologies. The final identification of the gene and regulatory sequence responsible for the phenotype is a key restriction of gene discovery based on genetic research. Omics research is quite trustworthy for identifying plausible candidates that are responsible for elucidating the biological function or process that affects the gene effect.18
Data from proteomics, transcriptomics and metabolomics can be evaluated for parental lines that offer significant support by providing knowledge about the genes in the target region. If the goal is a QTL related to heat tolerance, transcriptomics will reveal the local genes that are up- or down-regulated in response to heat stress. A segregating population can also be directly mapped using metabolite, protein, and transcript profiles to learn more about the loci that regulate gene expression, protein modification, or amounts of a particular secondary metabolite. The expression (eQTL), protein (pQTL), or metabolite (mQTL) QTLs are those linked to those attributes. The mapping pattern of eQTLs that regulate complex traits can be used to identify key regulatory regions for molecular breeding methods. An affordable alternative to costly eQTL and mQTL studies is transcript or metabolite profiling of selected recombinant lines with divergent phenotype from the overall mapping population.19-20 In order to identify the gene and characterise it afterwards, accurate phenotyping of the desired trait is crucial.
High-throughput phenotyping, often known as “phenomics,” has become a very busy study subject. It is never easy to analyse complicated traits using molecular breeding. This has been backed up by advancements in phenotypic data modelling and statistical methods that have been derived from field and controlled environment investigations.21 Omics assisted in bridging the gap between traditional phenotypic and genetic studies and physiological and molecular information. The creation of such an interaction model can aid in the connection between agronomy and gene structure and offer details on the selection targets that offer the most genetic benefit. Linkage drag or the impact of adverse alleles at closely spaced loci can sometimes make it difficult for breeders to successfully introduce certain wild alleles into adapted germplasm. Designing the most effective crossing and selection approach requires an understanding of the recombination behavior at and near the target locus. It helps to overcome linkage drag. Genomic tools can be used in this successfully. Analysing the genotype/environment link is made possible by whole genomes and related omics data. Therefore, it is clear that plant genomics offers a platform for examining and comprehending the genetic and molecular foundation of all biological activities in plants those are important to the species. With the aid of genomics, it is possible to evaluate hundreds of genes concurrently and comprehend complex crop properties like yield.22 Also, it closes the genetic and phenotypic gap and aids in understanding a gene’s interactions with other genes and genetic networks, which can influence a gene’s activity in addition to its isolated effects.23 Marker-assisted breeding will gradually transform into “genomics-assisted breeding” for crop enhancement with genomic techniques.24
Nowadays, proteome analysis is becoming popular among plant researchers to examine developmental changes in plants and to check the influence of environmental stimuli on protein patterns.25 Proteome analysis helps in better understanding of complex cellular events of plants. Proteome analyses are typically performed on mutants to assess how they react to various hormonal interventions, dietary considerations, and photosynthetic characteristics. Different unknown novel genes coding for enzymes of different metabolic pathways can be identified by proteome analysis.26 The plant proteome can undergo significant alterations as a result of abiotic stressors, which appear as the up- or down-regulation of proteins or as post-translational modifications to those proteins.27
All of an organism’s metabolites are identified and measured at one time using the technology known as metabolomics, which deals with comprehensive analysis.28 Metabolomics offers the capability of genotype and phenotypic identification and differentiation based on metabolic levels. Taking together, the genomics revolution is creating a revolution in plant breeding that cannot be ignored. These modern approaches are fundamentally important in addressing the need for enhancing agricultural productivity and sustainability, and to put them to use for the public good.
Genomic approaches
Heat tolerance is not a single step rather a complex phenomenon. Different types of abiotic stress can create a protective mechanism, so that a plant becomes tolerant or adapted to it. Signal recognition and transduction are two consecutive steps in the stress response, which ultimately result in to molecular, biochemical, and physiological reactions.29-30 With the development of omics techniques and strong bioinformatics expertise, the study of plant responses to abiotic stressors may be approached broadly. So far, 94,000 to 96,000 genes of wheat have been sequenced through shot gun sequencing. Additionally, two-thirds of these genes were assigned to the three hexaploid wheat genomes (A, B, and D).31 With the use of these extensive genetic resources and functional genomics methods, we can recognize and comprehend the cellular networks that are associated in signal perception, transduction, and defense response. These approaches help us to manipulate the genes and ultimately the expression of these stress related genes.
Demonstration of the genetic and molecular basis of all biological processes relevant to the particular species of plants is the most fundamental objective of plant genomics. Currently, the genomic programme produces huge numbers of data which includes gene sequence information, mutations, markers, maps, functional discoveries, etc. that need processing, storage and distribution towards the research community. Bioinformatics is the answer to all these necessities which is growing very fast nowadays.32
In plant heat tolerance, a crucial role is played by different heat shock proteins (HSPs) as well as heat shock factors (HSF). In response to heat and other abiotic stimuli, many scientific groups functionally characterized wheat’s heat shock proteins and components. In one study, out of 53 HSFs which was reported to date in wheat, few TaHSFs including one identified (TaHsfA2d) was selected for the expression profiling.33These HSFs’ diverse spectrum of expression in many tissues and differences in their expression in response to heat stress and different other abiotic stimuli demonstrated their usefulness as candidate genes. Performing position-Specific Scoring Matrix (PSSMs) of known HSP domains and sequence homology, Kumar et al., 2020 were able to identify 753 TaHSPs.34 They comprise 84 TaHSP100, 169 TaSHSP, 273 TaHSP40, 95 TaHSP60, 114 TaHSP70, and 18 TaHSP90. Apart from the role in high-temperature stress, the role of TaHSPs in seed development was confirmed by gene structure, gene expression and phylogenetic relationship.
Caseinolytic Protease B (CLPB) proteins are essential for organisms throughout various developmental phases of heat stress. These are chaperone proteins of high molecular weight. Caseinolytic Protease B (CLPB) homologous proteins, such as Heat Shock Protein 101 (HSP101), are crucial for plant development and heat adaptation. This function is highly conserved among different species. Some earlier studies on this group have identified HSP101/CLPB copies in wheat. A clear map of the total gene family in genome is unknown till date.35-37 For a genome-wide analysis, 13 members of the TaCLPB gene family were chosen, and their expression patterns were examined in diverse tissues, developmental stages, and stress settings.38 Detail characterization reveals its subcellular localization and its functional regulations. These genes had been successfully cloned and their homoeologous expression patterns were studied under moderate and extremely high temperatures in four wheat genotypes. It is possible to improve wheat’s thermotolerance by focusing on one or more TaCLPB genes.
Monomeric proteins with a size range of 12-42 KDa are known as small heat shock proteins (sHSPs).39 When stressed by high temperatures, plants have a lot of them. These genes are generally encoded by nuclear genes. Their localization is distributed from cytoplasm/ Nucleus to ER, mitochondria, chloroplasts, etc.Hsp26 proteins that are localised in the chloroplast interact with Photosystem II, the part of the photosynthetic mechanism that is most sensitive to temperature.40-41 It guards the photosynthetic apparatus during heat stress. For this, a complete sequence based characterization is needed for this gene. In a study, four members of small heat shock proteins i.e. basically chloroplast-localized sHSP were isolated and characterized. The establishment of thermotolerance and prevention of irreversible accumulation of misfolded proteins are known to be facilitated by small heat shock proteins. sHSPs that are localised in the chloroplast shield the photosynthetic apparatus against high temperature stress. To find new alleles in this family, two high throughput TILLING (Targeting Induced Local Lesions In Genomes) techniques (in vivo and in silico) were used in this study.42 The ExpVIP, a customizable RNA-seq Data Analysis and Visualization platform, has been used to analyse the modulation of the newly discovered genes within the sHsp26 family in vivo. The scope of abiotic stress analysis has been widened by this study, which also demonstrates that these genes are involved in the plant body’s reaction to heat stress. Additionally, fifty TILLING mutant lines were identified in this study. A collection of KASP (Kompetitive Allele Specific PCR) markers that may be used with high throughput genotyping techniques was also designed to track the particular mutations in the continuing backcrosses.
The function of late embryogenesis abundant (LEA) protein gene family to minimize cell damage is well known. This group of functional protein effectively protect cells under abiotic stress conditions too.43-44This protein (LEA) is generally induced under various abiotic stresses and can be accumulated during late embryonic development.“Chinese Spring” wheat (Triticum aestivum) was chosen for first important survey on the LEA gene family.45 Researchers have discovered 179 TaLEA genes in this investigation, which have been divided into eight groups. Their findings demonstrated that the majority of TaLEA genes contained ABA-responsive elements (ABREs), and numerous cis-acting elements in the promoter region that are linked to the stress response and induced in response to ABA and abiotic stress. To determine the protective role of these genes, they inserted 8 genes, one for each group, into yeast and E. coli. Their result established the fact that these genes have a function in heat and salt stress. So, future functional investigations with an objective to improve the stress tolerance of wheat, LEA genes can be targeted as robust candidate genes.
Transcription factors (TFs) and the cis regulatory sequences they are associated with act as molecular regulators of gene expression, controlling the timing and location of gene expression, particularly under stressful conditions.46 Furthermore, wheat transcription factor databases like “PlantTFDB” (see: http://planttfdb v1.cbi.pku.edu.cn:9010/web/index.php?sp=ta) and “Database of Wheat Transcription Factor” (see: http:/wwwappli.nantes.inra.fr:8180/wDBFT/) would be useful tools for the scientists to investigate the role of Transcription factorss in stress response.
Based on conserved gene ontology, recent wheat genome sequencing has also shown the role of transcription factors under a variety of environmental situations in a number of model crops.46-50 Cumulatively 191 bZIP transcription factors from wheat has been discovered through homology searches.51 After phylogenetic analysis of wheat bZIP proteins and gene ontology enrichment analysis, one of the members named TabZIP was selected for expression analysis in different abiotic stress including heat stress. In varieties of abiotic stress conditions differential expression of TabZIP was established by gene expression studies which suggest the obvious role of bZIP in stress bypassing mechanisms. These conditions include heat, salinity and dehydration etc.Transgenic Arabidopsis expressing TabZIP show high tolerance against heat and other abiotic stress.
Transcriptomics approach
As heat stress is associated with different physiological and biochemical events and multiple genes are involved in this process, complete gene expression profiling can provide much better information than the study of a single gene. The recognition of several genes related to heat tolerance is now possible due to the use of contemporary techniques like microarrays for genome-wide expression profiling, subtraction libraries, expressed sequence tags (ESTs) from cDNA libraries, cDNA-AFLP, RNA-seq, serial analysis of gene expression (SAGE), and whole transcriptome sequencing.52-56 RNA-seq is superior to microarray as it is capable of detecting novel transcripts, gene knowledge is not essential, and offers opportunities to investigate noncoding RNAs.In wheat, like many other crops, short and lengthy heat treatments can cause the up- or down-regulation of several genes, including transcription factors, HSPs, phytohormone production and signaling specially calcium, and sugar signal pathways.57-60 The up-regulation of seven heat responsive genes in durum wheat under heat treatment was also reported using cDNA-AFLP based transcriptome profiling.61 miRNA regulated differential gene expression in heat stress was also tested by different research groups. Using solexa high-throughput sequencing in small RNAs of wheat, the existence of 9 putatively heat-responsive miRNA was established. After heat treatment, 8 miRNAs, specifically miR156, miR159, miR160, miR166, miR168, miR169, miR827, and miR2005, were up-regulated while miR172 expression was noticeably reduced.62 Expression of 79 miRNAs including miR398, miR528 and miR156, which are important for the activation of signal transduction pathways and flowering, are down regulated due to heat within 1 day of heat treatment.63Another study found that the wheat cultivar HD2985 that had been heat treated and the control had, respectively, 1052 and 902 mature miRNA sequences. 37 new miRNAs were discovered utilising various pipeline and plant-specific criteria, and six of them were confirmed to be heat-responsive using qRT-PCR.64
On the other hand, transcriptome profiling was done during early and late grain filling stage (14 and 30 days post anthesis) in different heat susceptible and tolerant genotypes. In response to heat stress, up-regulation of bunch of genes comprising 6-phosphogluconate dehydrogenase, peptidylprolyl isomerase, FtsH protease,S6 RPS6-2 ribosomal protein, heat shock cognate-70, plasma membrane proton ATPase, methionine synthase, RuBisCO activase B, HMW-glutenin and cytochrome C (class I), were observed in heat stress tolerant genotypes like Fang-60 and in case of susceptible genotypes like Gregory and banks these genes are down regulated.65 Plant molecular biologists have studied the temperature effect on the embryos of wheat seeds using the transcriptomics approach. Researchers demonstrated the upregulation of a wheat homolog of MOTHER OF FT AND TFL1 (MFT) following their physiological development in dormant seeds, i.e. grown at somewhat lower temperature. Additionally, they demonstrated their co-localization with the QPhs.ocs-3A QTL, which controls seed dormancy.66 The physiological and molecular effects of incorporating a foreign chromosomal segment (7DL) from Agropyron elongatum, a wild relative of wheat, into Triticum aestivum (cultivated wheat) were investigated. Higher root and shoot biomass along with improved water stress adaptation was observed in wheat translocation line. Two candidate genes were identified by transcriptome analysis that are associated with root development. Using single-feature polymorphism analysis, the candidate genes were found to be located on the translocated region of chromosome 7DL.67
Using deep RNA sequencing technology another research group showed out of 4,375 wheat transcription factors, 1,328 were responsive to stress treatments. Involvement of HSPs and DREBs was also reported by them in response to heat, drought and their combination.68
Proteomics approach
To get the most information out of plant tissue expression profiling, it is crucial to recognize the protein constituents. Analysis of protein, genomic sequence, and biological activity can all be used to establish a direct connection. With the analysis of proteome, we not only acquire detailed knowledge about the protein but also other aspects like expression level of the protein, post-translational modifications, and interactions, etc. For this, a sensitive and accurate proteome analysis technique is becoming a useful tool for recognizing the proteins associated in biotic and abiotic stress. The varying amounts of different enzymes and/or proteins of numerous metabolic pathways reflect global variations in the expression of plant genes during growth, development, and exposure to altered environments.69
By proteome analysis of a tolerant wheat line key enzymes or metabolic intermediates can be identified. These may be targeted later to design heat-tolerant wheat varieties. Matrix-assisted laser desorption/ionization-time of flight mass spectrometry (MALDI-TOF-MS) and two-dimensional electrophoresis (2-DE) were employed to examine changes in protein expression during heat stress. Researchers discovered 43 proteins with differential expression, including ATP synthase, heat shock proteins, and numerous defense proteins, as well as enzymes involved in carbohydrate metabolism like glucose-1-phosphate adenyl-transferase and granule-bound starch synthase.70 Enzymes that are responsible for carbohydrate metabolism may play a role in the starch degradation in susceptible genotypes, or the formation of starch in tolerant genotypes during the grain filling stage. Although wheat has been found to express numerous heat shock proteins (HSPs) under high temperature conditions, their specific functions in response to heat stress are not yet fully comprehended. The low molecular weight HSPs have shown differential expression in two wheat cultivars, the heat-tolerant (cv. Fang) and the heat-susceptible (cv. Wyuna), leading to the discovery of a number of distinct isoforms. High molecular weight HSPs like HSP70 and HSP90 as well as low molecular weight HSPs like HSP26 are all heat-induced HSPs in wheat. HSP70 is the one which is specifically connected to thermotolerance.71 On the other hand, heat stress has a strong inducible effect on the chloroplast HSP (HSP26) in practically all vegetative and generative tissues of wheat.72 Another research group found that under heat stress, the levels of antioxidants (2-Cys peroxiredoxin BAS1), heat shock protein (HSP70), and proteins engaged in photosynthesis (such as Rubisco activase, sedoheptulase-bis-phosphatase, and fructose bisphosphate aldolase, which are involved in the generation of RUBP), proteins involved in signal transduction (BRI-1KD interacting protein 114) increased. The levels of the enzymes involved in nitrogen metabolism decrease in resistant cultivars as a result of heat stress during the grain filling stage.73
There are several other heat sensitive proteins in wheat. Dough extensibility and elasticity are related to gliadins and glutenin proteins. According to a proteomics analysis, exposure to heat stress resulted in a notable increase in gliadin levels, while the synthesis of glutenins remained unaffected. This suggests that the accumulation of gliadins is more sensitive to heat than that of glutenins.74 Several research groups also investigated how albumin and gliadin were affected by high-temperature stress (32 °C).75
As a result of TaHsfA6f, a member of the A6 subclass of heat shock transcription factors, being overexpressed in green organs of wheat plants, heat shock proteins (HSPs) and several other heat stress protection genes, including Golgi Anti-Apoptotic Protein (GAAP) and the large isoform of Rubisco activase, were upregulated.76 Using comparative proteome study of flag leaves using the LC-MS/MS technology, about 258 heat-responsive proteins (HRPs), including several proteins involved in chlorophyll production and carbon fixation, were discovered under heat stress. In addition to HSP100, HSP70, HSP 90, and HSP20, a noticeably increased accumulation of glutathione-S-transferase (GST) and thioredoxin (Trx) in response to heat demonstrates their significance in the regulation of redox reactions.77 Identification of 19 differentially expressed proteins including HSP70, Oxygen evolving enhancer protein, calmodulin, hypothetical protein, stress predicted protein, Rubisco, Rubiscoactivase, signalling molecule (CDPK) in control and heat shock (HS) treated samples at a milky-ripe stage of wheat using two-dimensional gel electrophoresis coupled with MALDITOF/MS, Nano-scale liquid chromatographic tandem mass spectrometry (nLC/MS) and MS2 was done.78 14 protein spots that were differently regulated in wheat seedlings were identified by 2-DE combined with MALDI TOF/TOF. Three proteins, namely PEP carboxylase 1, ConA and Rubisco activase A as well as the overexpression of the catalase and cytochrome b6f complex in tolerant lines, were differentially regulated and demonstrated to be involved in the heat defense mechanism.79
Metabolomics approach
Metabolomics is a field of study that focuses on the recognition and measurement of metabolites present within a biological system, such as a cell, tissue, or organism. Metabolites are small molecules that are engaged in various biochemical processes, including energy metabolism, signaling, and regulation of gene expression. By analyzing the metabolome, researchers can gain insights into the physiological and biochemical state of a biological system, and how it responds to various environmental and genetic factors. Metabolomics has applications in a wide range of fields, including medicine, agriculture, and environmental science.
Plants face different abiotic stresses from time to time. They may acclimatize to challenging environmental conditions by modifying the composition and concentration of metabolites. Global metabolite profiling is essential for understanding the precursors, intermediates, and end/final products of the metabolic pathways in any organism, including plants.80 Metabolomics became useful and important tool for the analysis of traits in response to herbicide also.81
Gas or liquid chromatography-mass spectrometry (GC-MS and LC-MS), Nuclear magnetic resonance (NMR) and capillary electrophoresis-mass spectrometry (CE-MS) are frequently utilized in plant sciences.82 Numerous bioinformatics tools and statistical models have been developed for interactive analysis of the metabolome.83 Durum wheat showed an increase in certain metabolites such as sucrose and glycerol under heat stress, which can impact the nutritional quality of the grain even with short-term exposure to high temperatures.84
Transgenic approach
Plant biotechnology techniques offer a faster and more appealing alternative to traditional breeding and marker-assisted selection programs for introducing a small number of genes directly into plants to overcome various abiotic stress factors, including heat. Currently, researchers are exploring the integration of one or more genes that code for proteins involved in biochemical pathways or signaling pathways endpoints, under the effect of a constitutively active promoter.Stress tolerance of transgenic plants need mechanisms of highly expressed proteins related to osmotic adjustments. To combat stress-related issues, a saturation in membrane lipids, removal of reactive oxygen species (ROS), photosynthetic processes, polyamine generation, and protein biosynthesis are used. 85 By overexpressing stress-responsive genes and heat shock transcription factors, stress tolerance in transgenic plants has been accomplished. 86-88 It is quite evident that molecular control of gene expression, regulation of their spatial and temporal expression, specifically during stress is somehow dependent on transcription factors (TFs) and the cis-regulatory sequences they correspond to.89 The analysis of Ae. Tauschii revealed the presence of 1,489 transcription factors (TFs) categorized into 56 families, with a significant proportion of them belonging to the MYB-related TFs. These MYB-related TFs are involved in stress-tolerance.44
Different scientific groups expressed various transcription factors in wheat to achieve stress tolerance including heat. Heat stress is widely recognized as a cause of protein denaturation and aggregation, as well as membrane damage, which can result in changes to metabolic fluxes. The plastidal protein synthesis elongation factor (EF-Tu) gene from maize was used to create transgenic wheat.90 These plants showed less thermal leaf protein aggregation as compared to non-transgenic plants, less heat stress-related harm to photosynthetic membranes (thylakoids), and a higher rate of CO2 fixation. The overexpression of TaNAC2L, a transcription factor from wheat, was also shown in another study to improve heat tolerance by triggering the expression of temperature-related genes in Arabidopsis.91 A transgenic line of Arabidopsis containing another transcription factor, TaWRKY33 of wheat is tolerant to heat stress than the control plant.92 Cloning and expression of transcription factors became a common tool to achieve heat tolerance in wheat. Cloning of the transcription factor ArabidopsisWRKY30 (AtWRKY30) and expression of it in wheat demonstrate higher biomass, growth, chlorophyll content, gas-exchange characteristics, proline, relative water content, soluble sugar content, soluble proteins content and antioxidant enzyme activities than wild plants.93 In comparison to wild plants, the AtWRKY30-overexpressing wheat plants have relatively high levels of malondialdehyde content, electrolyte leakage and hydrogen peroxide.
Numerous research papers have noted the role of Hsp proteins in safeguarding cells from heat damage, as previously discussed. According to numerous study, during heat stress, Hsp proteins work as molecular chaperones to maintain the natural structure and activity of cell proteins. Of the five families of conserved Hsps (Hsp60, Hsp70, Hsp90, Hsp100, and sHsp), small heat shock proteins (sHsps) are highly abundant in plants, and their expression can increase by as much as 200-fold when exposed to heat.
The causal involvement of HSPs in plant heat tolerance is confirmed by experimental results from transgenic, reverse genetics, including down-regulating the levels of HSPs by antisense and RNAi method and mutation approaches in cereal and non-cereal species.94 Mutant varieties of A. thaliana and Z. mays that expressed lower levels of Hsp100 were found to exhibit reduced basal and induced heat tolerance.95-96 Heat shock proteins that interact with regulatory HSF proteins positively influence the transcription of hsp genes.97 Numerous numbers of hsp genes and heat shock transcription factors have been cloned and characterized.98 The Hsp90 gene, which spans 2,323 base pairs and has an open reading frame that is between 62 and 2,164 base pairs long and codes for 700 amino acids, was cloned and characterised from the wheat cultivar C-306.qRT PCR expression analysis revealed that in the C-306 cultivar, the level of transcripts of the hsp90 gene increased by 1.5-fold, 1.2-fold, and 2.5-fold in the root during the pollination, milky dough, and seed hardening stages, respectively. Meanwhile, the expression levels in the flag leaf showed a 4.5-fold, 4.3-fold, and 6.5-fold increase during the same stages, respectively. Upon subjecting C-306 and PBW343 (a susceptible cultivar) to varying levels of heat shock, a noticeable difference was observed in the fold expression of HSP90.So, Hsp90 can be a potential target to develop a heat-tolerant wheat variety.99 A lot of Hsp gene expression study shows its potential to counter heat stress. In another study, researchers expressed the hsp26 gene of T. aestivum in trans host Arabidopsis. When compared to their wild-type counterparts, engineered plants showed elevated levels of tolerance to ongoing high temperatures.100 Maize HSP101 is essential for primary root growth and induced and basal thermotolerance.101Alterations to the expression of a small heat shock protein gene, hsp17.7, in carrots have been shown to result in changes to the plant’s thermo tolerance levels, both increases and decreases.102
Transcription factors (HSFs) for heat stress, the last elements of signal transduction pathways, including their own activation, mediate the activation of genes in response to heat stress. The modification of the effects of the heat shock transcription factor is also very much crucial for heat tolerance. When expressed in Arabidopsis, TaHsfA2d, a heat shock transcription factor from wheat that is preferentially expressed in seeds, exhibits a high degree of thermo tolerance.103 Studies shown overexpression of a heat shock factor, TaHsfC2a improved thermo tolerance in wheat through an ABA-mediated regulatory pathway.104
It is typical for plants to accumulate different osmolytes in response to abiotic stress. To safeguard cellular structures from the damaging effects of elevated temperatures and to control osmotic activities, these osmolytes are essential for membrane stability, maintaining cellular water balance and buffering the cellular redox potential.105 Various osmolytes, including amino acids (such as proline), polyamines (such as putrescine), quaternary ammonium compounds (such as glycine-betaine), sugars (such as fructans, mannitol, sorbitol, and trehalose), and sugar alcohols (such as polyols), accumulate in plants as a response to osmotic stress. So, it is an obvious belief of plant biotechnologists that heat tolerant transgenic plants can be elevated by changing the osmolytes level in plants. Production of glycine-betaine can be increased by transforming the BADH gene in plants.It is thought to be a potentially effective strategy for growing plants that can withstand heat.106 The transgenic wheat line T6 showed an improvement in heat tolerance through over-accumulation of glycine-betaine, which was linked to improved osmotic adjustment and improved antioxidant defence mechanisms using antioxidants and antioxidative enzymes.107 Similarly, reactive oxygen species have a close attachment with heat stress. Overproduction of ROS during heat stress cause oxidation of biomolecules like- protein, lipid, carbohydrate, nucleic acid, photosynthetic pigments which ultimately jeopardize the normal cellular functions.108 By over expressing the enzymes which act as ROS scavenger, heat tolerance can be achieved. Transgenic Arabidopsis plants that overexpressed the hvapx1 gene, responsible for encoding peroxisomal ascorbate peroxidase from barley, demonstrated increased heat tolerance compared to their wild-type counterparts.109 Additionally, it has been noted that plants are protected against the effects of high-temperature stress when Cu/Zn superoxide dismutase is overexpressed.110
For continuous photosynthesis and respiratory function, cellular membrane function must be maintained under high temperature stress.111 So, heat tolerant plants can be produced by altering membrane fluidity.By turning off the gene for chloroplast omega-3 fatty acid desaturase, transgenic tobacco plants were able to produce modified chloroplast membranes that contained more dienoic fatty acids and less trienoic fatty acids than their wild-type counterparts. This resulted in higher rate of photosynthesis and improved growth under high-temperature conditions.112
Apart from the proteins mentioned above some other heat responsive gene, as well as proteins can be used as a tool to counter heat stress in plants. Experimental data suggested that TaLTP3, a wheat lipid transfer protein 3, could improve Arabidopsis’s seedling-stage baseline thermotolerance and oxidative stress resistance.113 Singh and Khurana did molecular and functional characterization of wheat B2 protein and showed it is overexpression can increase heat tolerance in Arabidopsis.114 A heat-responsive gene’s (TaGASR1) isolation and characterisation from wheat was performed, and it has been demonstrated that overexpressing this gene in wheat and Arabidopsis increases tolerance to heat stress and oxidative stress.115
When transgenic Arabidopsis plants expressed TaOEP16-2-5B, a gene encoding a wheat plastid outer envelope protein, they displayed increased tolerance to heat and drought stress.116 In another study, this same scientific group overexpressed the wheat ferritin gene in TaFER-5B and proved how it controls different abiotic stress including heat by the ROS scavenging pathway.117 Transcriptomics study led to the discovery of the gene TaPEPKR2, which belongs to the phosphoenolpyruvate carboxylase kinase family. It is located on chromosome 5B. Transgenic lines with this gene showed improved resistance to heat and dehydration stress.118
Epigenetic control on heat tolerance
In modern times, the term epigenetics has become commonplace. It refers to changes in gene expression and activity that do not involve modifications to the underlying DNA sequence.DNA methylation, histone modification, and noncoding RNA are responsible for it. Epigenetic regulations on heat tolerance are gaining interest day by day.119-120 Studies have shown that the histone acetyltransferase GCN5 is essential for keeping Arabidopsis thermotolerant. Specifically, it achieves this by acetylating H3K9 and H3K14 on heat shock factor A3 (HSFA3) and UV-HYPERSENSITIVE6 (UVH6) during periods of elevated heat stress. When subjected to high temperatures, the TaGCN5 gene in wheat is upregulated and acts similarly to Arabidopsis’ GCN5 histone acetyltransferase gene. The relaxation of chromatin structure is linked to the activation of transcription. Consequently, the impact of heat stress on the promoters of six widely recognized genes (TaHSF1, TaHSF4, TaMBF1c, TaHSP17.4, TaHSP26, and TaHSP101) was investigated by analyzing the levels of H3K9 and H3K14 acetylation. CHIP assay suggested acetylation was increased in five genes except for HSP26 after heat treatment.121 The effect of temperature has a huge effect on gene expression. A recent study that surveyed the hexaploid wheat genome revealed minimal differences in methylation patterns between plants grown under two different temperatures (12 °C and 27 °C). Methylation was only occasionally linked to minute alterations in gene expression.120
Recent studies indicate that a significant part of the genome is transcribed, but only a small fraction of it actually codes for proteins. The huge amount of transcripts that do not translate into protein is known as non-protein coding RNA (npcRNAs). Non-coding RNAs (ncRNAs) can be classified into two main types: housekeeping ncRNAs (including transfer and ribosomal RNAs) and regulatory ncRNAs or riboregulators. The regulatory ncRNAs can be further classified based on their length into short regulatory npcRNAs (less than 300 base pairs long), which include microRNA, siRNA, and piwi-RNA, and long regulatory npcRNAs (more than 300 base pairs long).62,122-124 37 novel miRNA from wheat was recognized and found six of them as heat-responsive.62 Upon subjecting durum wheat cv Cappelli and Ofanto to heat treatment, researchers observed differential expression of several pre-miRNAs. Specifically, they identified 12 pre-miRNAs in cv Cappelli and 25 pre-miRNAs in Ofanto that showed altered expression levels.124 A team of scientists conducted an independent study and found that wheat plants subjected to 2 hours of heat stress exhibited a downregulation of TamiR159. Interestingly, when rice lines overexpressing TamiR159 were exposed to heat stress, they displayed a greater sensitivity to the stress compared to the wild type. This shows that a heat stress-related signalling pathway may be involved in the downregulation of TamiR159 in wheat during heat stress.125
Researchers utilized small RNAs and degradome analysis (which involves analyzing RNA ends in parallel) on leaf tissues obtained from both control and heat-stressed wheat plants. By doing so, they have identified and validated miRNAs that were regulated by heat stress, as well as their corresponding target genes that are involved with thermo tolerance. These findings could help shed light on the molecular mechanisms involved in heat stress response in wheat plants.126 These studies showed how miRNA is highly altered in heat stress and again returns to anormal level after removal of heat stress. Through degradome sequencing, researchers found that the target genes of several miRNAs, including miRNA156, miR159, miR166, and miR398, are conserved across various cereal crops, including wheat, as well as dicot plants. This suggests that these miRNAs could have important functional roles that are conserved across numerous plant species.
Conclusion
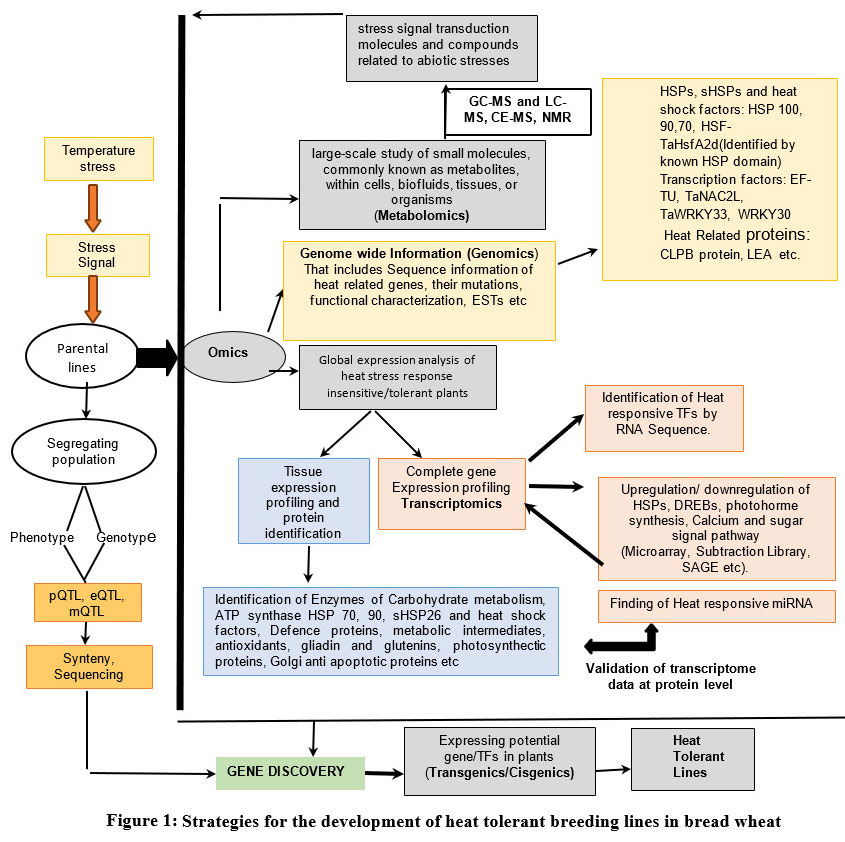
It is challenging to measure the polygenic trait of heat tolerance. Although there has never been a straightforward technique to choose heat-tolerant plants, several characteristics, such as membrane thermo-stability and canopy temperature depression, seem to be good predictors of plant heat tolerance.127 A thorough and integrated approach combining physiology, genetics, genomics, and genetic engineering is needed to unravel the intricate processes of heat stress, which are controlled by numerous genes. Advancements in the field of omics have provided researchers with new opportunities to explore various aspects of abiotic stress tolerance in wheat. Omics-based approaches have aided in a deeper comprehension of the mechanisms behind stress response and tolerance in wheat plants by generating enormous amounts of data and information. Genomic, transcriptomic, and proteomic research on wheat has advanced, but metabolomics and phenomics, two other important omic branches, are still lagging behind. Understanding the genetic basis of heat stress tolerance in wheat has tremendously benefited from various mapping techniques and genetic research.128 Molecular markers identified in these studies as being linked to heat tolerance may be applied to MAS. Molecular interaction between different biomolecules is revealed through interactome network studies. Additionally, it advances our understanding of the genotype-phenotype link. By combining the insights gained from transcriptomics, proteomics, genomics, and metabolomics studies, researchers can obtain a more holistic understanding of the gene networks involved in heat tolerance in wheat. It is conceivable to discover significant genes and pathways that might be targeted to improve wheat’s ability to tolerate heat stress and obtain a full grasp of the molecular mechanisms behind stress response by employing such an integrated technique. The convergent amalgamation of omics as depicted in Fig1 is probably the most promising way for developing wheat cultivars with heat tolerance.
 |
Figure 1: Strategies for the development of heat tolerant breeding lines in bread wheat. |
{kind=link}
Acknowledgment
Authors thankfully acknowledge Dr. Md Nasim Ali, Head of the Department, Agricultural Biotechnology and all lab mates of BCKV for providing constant support and new ideas for research.
Conflict of Interests
The authors have no relevant financial or non-financial interests to disclose
Funding sources
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript
References:
- CGIAR (2022) Research program on wheat. Accessed: 12 January 2022; https://cgspace.cgiar.org/bitstream/handle/10568/89829/WHEATWeb.pdf?sequence=4&isAllowed=y
- Barros V. R., Field C. B., Dokken D. J., Mastrandrea M. D., Mach K. J., Bilir T. E., Chatterjee M., Ebi K. L., Estrada Y. O., Genova R. C., Girma B., Kissel E. S., Levy A. N., MacCracken S., Mastrandrea P. R., White L. L. (eds.). Climate change: impacts adaptation and vulnerability. Part B: regional aspects. Cambridge: IPCC; 2014; Cambridge University Press, Cambridge: 688.
- Driedonks N., Rieu I., Vriezen W. H. Breeding for plant heat tolerance at vegetative and reproductive stages. Plant reprod. 2016; 29(1): 67-79.
- Young L.W., Wilen R.W., Bonham-Smith P.C. High temperature stress of Brassica napus during flowering reduces micro-and mega gametophyte fertility induces fruit abortion and disrupts reproduction. Exp.Bot. 2004; 55(396): 485–495.
- Zinn K. E., Tunc-Ozdemir M., Harper J. F. Temperature stress and plant sexual reproduction: uncovering the weakest links. Exp.Bot. 2010; 61(7): 1959–1968.
- Vollenweider P., Günthardt-Goerg M.S. Diagnosis of abiotic and biotic stress factors using the visible symptoms in foliage. Pollut. 2005; 137(3) :455-465.
- Barnabás B., Jäger K., Fehér A. The effect of drought and heat stress on reproductive processes in cereals. Plant, Cell & Environment. 2008; 31(1): 1–38.
- Hays D., Mason E., Hwa Do J., Menz M., Reynolds M. Expression quantitative trait loci mapping heat tolerance during reproductive development in wheat ( aestivum). In: Buck, H.T., Nisi, J.E., Salomo’n, N. (Eds.). Wheat production in stressed environments. Amsterdam: Springer; 2007: 373–382.
- Rane J., Shoran J., Nagarajan S. Heat stress environments and impact on wheat productivity in India: Guestimate of losses. Indian Wheat News Letter. 2000; 6:5–6.
- Enghiad A. D., Countryman A. M., Thilmany D. D. An Overview of global wheat market fundamentals in an era of climate concerns. J. Agron. 2017; 19: 1-15.
- Umarani E., Hemalatha V., Saritha A., Ramanjaneyulu A.V. Impact of high temperature stress in Rice. J.Econ.Plants.2020; 7(3): 108-110.
- Manavalan L. P., Guttikonda S. K., Phan Tran L. S. , Nguyen H. T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009; 50:1260–1276
- Setia R. C., Setia N. The ‘omics’ technologies and crop improvement. In: Setia R. C., Harash N., Setia N. (Eds.) Crop improvement: strategies and applications. New Delhi: International Publishing House Pvt. Ltd; 2008; 1–17.
- Xu Y. F., An D. G., Liu D. C., Zhang A M., Xu H X., Li B. Mapping QTLs with epistatic effects and QTL × treatment interactions for salt tolerance at seedling stage of wheat. Euphytica. 2012; 186: 233–245.
- Miflin B. Crop improvement in the 21st century. Exp. Bot. 2000; 51: 1-8.
- Paleman J. D., Vander Voort J. R. Breeding by design. Trends Plant Sci. 2003; 8: 330-334.
- Ebeed H. T. Omics Approaches for Developing Abiotic Stress Tolerance in Wheat. In: Hasanuzzaman M., Nahar K., Hossain M. (Eds.), Wheat Production in Changing Environments. Singapore: Springer; 2019; 443-463.
- Langridge P., Fleury D. Making the most of ‘omics’ for crop breeding.Trends Biotechnol. 2011; 29(1): 33-40.
- Fernie A.R., Schauer N. Metabolomics-assisted breeding: a viable option for crop improvement? Trends Genet. 2009; 25:39–48.
- Marino R., Ponnaiah M., Krajewski P., Frova C., Gianfranceschi L., Pè M. E., Sari-Gorla M. Addressing drought tolerance in maize by transcriptional profiling and mapping. Genet. and Genom. 2009; 281(2): 63– 217.
- Hammer G., Cooper M., Tardieu F., Welch S., Walsh B., Eeuwijk F., Chapman S., Podlich D. Models for navigating biological complexity in breeding improved crop plants. Trends Plant Sci. 2006; 11: 587–593.
- Struik P. C., Cassman K. G., Koornneef M. A dialogue on interdisciplinary collaboration to bridge the gap between plant genomics and crop sciences. In: Spiertz J. H. J., Struik P. C., VanLaar H. H. (Eds.), Scale and Complexity in Plant Systems Research: Gene-Plant-Crop Relations. The Netherlands: Springer; 2007; 317-326.
- Campos-de Q. H. Plant genomics: an overview. Res. 2006; 35: 385-399.
- Varshney R. K., Graner A., Sorrels M. E. Genomics-assisted breeding for crop improvement. Trends Plant Sci. 2005; 10: 621-630.
- Setia R. C., Setia N. The ‘omics’ technologies and crop improvement. In: Setia R. C., Harash N., Setia N (Eds.), Crop improvement: strategies and applications. New Delhi: International Publishing House Pvt. Ltd; 2008; 1–17.
- Canovas F. M., Dumas-Gaudot E., Recorbet G., Jorrin J., Mock H. P., Rossignol M. Plant proteome analysis. Proteomics. 2004; 4: 285-298.
- Rampitsch C., Srinivasan M. The application of proteomics to plant biology: a review.J. Bot. 2006; 84: 883- 892.
- Fiehn O. Metabolomics — the link between genotypes and phenotypes. In: Functional Genomics (Eds. Town C.), Dordrecht: Springer; 2002; 155-171.
- Hall R. D., Vos C. H. R., Varhoeven H. A., Bino R. J. Metabolomics for the assessment of functional diversity and quality traits in plants. In: Vaidyanathan S., Harrigan G. G., Goodacre R (Eds.), Metabolome Analyses: Strategies for Systems Biology. New York: Springer; 2005; 31-44.
- Komatsu S., Yamamoto R., Nanjo Y., Mikami Y., Yunokawa H., Sakata K. A. comprehensive analysis of the soybean genes and proteins expressed under flooding stress using transcriptome and proteome techniques. Proteome Res. 2009; 8: 4766–4778.
- Brenchley R., Spannagl M., Pfeifer M., Barker G. L., D’Amore R., Allen A. M., McKenzie N., Kramer M., Kerhornou A., Bolser D., Kay S., Waite D., Trick M., Bancroft I., Gu Y., Huo N., Luo M. C., Sehgal S., Gill B., Kianian S., Anderson O., Kersey P., Dvorak J., McCombie W. R., Hall A., Mayer K. F., Edwards K. J., Bevan M. W., Hall N. Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature. 2012; 491: 705-710.
- Elanchezhian R. Application of Bioinformatics in Agriculture. In: Singh K. M. Meena M. S. (Eds.), ICT for agricultural development in changing climate (1st ). New Delhi: Narendra Publishing House. 2012; 163-179.
- Agarwal P., Khurana P. Functional characterization of HSFs from wheat in response to heat and other abiotic stress conditions. Integr.Genomics. 2019; 19: 497–513.
- Kumar A., Sharma S., Chunduri V., Kaur A., Kaur S., Malhotra N., Kumar A., Kapoor P., Kumari A., Kaur J., Sonah H., Garg M. Genome-wide Identification and Characterization of Heat Shock Protein Family Reveals Role in Development and Stress Conditions in Triticum aestivum Sci. Rep. 2020; 10: 7858.
- Wells D. R., Tanguay R. L., Le H., Gallie D. R. HSP101 functions as a specific translational regulatory protein whose activity is regulated by nutrient status. Genes Dev. 1998; 12: 3236–51.
- Campbell J. L., Klueva N. Y., Zheng H. G., Nieto-Sotelo J., Ho T.D., Nguyen H.T. Cloning of new members of heat shock protein HSP101 gene family in wheat (Triticumaestivum (L.) Moench) inducible by heat, dehydration, and ABA. Biochimica et Biophysica Acta (BBA)- Gene structure and Expression. 2001; 1517(2): 270-277.
- Muthusamy S. K., Dalal M., Chinnusamy V., Bansal K.C. Differential Regulation of Genes Coding for Organelle and Cytosolic ClpATPases under Biotic and Abiotic Stresses in Wheat. Plant Sci. 2016; 7: 929.
- Erdayani E., Nagarajan R., Grant N. P., Gill K. S. Genome-wide analysis of the HSP101/CLPB gene family for heat tolerance in hexaploid wheat. Rep. 2020; 10(1):1-17.
- Waters E.R. The evolution, function, structure, and expression of the plant sHSPs. Exp. Bot. 2013; 64: 391–403.
- Osteryoung K.W., Vierling E. Dynamics of small heat shock protein distribution within the chloroplasts of higher plants. Biol. Chem. 1994; 269: 28676–28682.
- Joshi C. P., Klueva N.Y., Morrow K. J., Nguyen H.T. Expression of a unique plastid-localized heat-shock protein is genetically linked to acquired thermo tolerance in wheat. Appl.Genet.1997; 95: 834–841.
- Comastri A., Janni M., Simmonds J., Uauy C., Pignone D., Nguyen H. T., Marmiroli N. Heat in Wheat: Exploit Reverse Genetic Techniques to Discover New Alleles Within the Triticum durum sHsp26 Family. Plant Sci. 2018; 9:1337.
- Debnath M., Pandey M., Bisen P. S. An omics approach to understand the plant abiotic stress. Omics. 2011; 15: 739–762.
- Hirayama T., Shinozaki K. Research on plant abiotic stress responses in the post-genome era: past, present and future. Plant J. 2010; 61: 1041–1052.
- Liu H., Xing M., Yang W., Mu X., Wang X., Lu F., Wang Y., Zhang L. Genome-wide identification of and functional insights into the late embryogenesis abundant (LEA) gene family in bread wheat (Triticum aestivum).Rep. 2019; 9: 13375.
- Garg B., Lata C., Prasad M. A study of the role of gene TaMYB2 and an associated SNP in dehydration tolerance in common wheat. Biol.Rep. 2011; 39: 10865-10871.
- Riechmann J. L., Heard J., Martin G., Reuber L., Jiang C., Keddie J., Adam L., Pineda O., Ratcliffe O. J., Samaha R., Creelman R., Pilgrim M., Broun P., Zhang J. Z., Ghandehari D., Sherman B. K., Yu G. Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science. 2000; 290: 2105-2110.
- Brenchley R., Spannagl M., Pfeifer M., Barker G. L., D’Amore R., Allen A. M., McKenzie N., Kramer M., Kerhornou A., Bolser D., Kay S., Waite D., Trick M., Bancroft I., Gu Y., Huo N., Luo M. C., Sehgal S., Gill B., Kianian S., Anderson O., Kersey P., Dvorak J., McCombie W. R., Hall A., Mayer K. F., Edwards K. J., Bevan M.W., Hall N. Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature. 2012; 491: 705-710.
- Ling H. Q., Zhao S., Liu D., Wang J., Sun H., Zhang C., Fan H., Li D., Dong L., Tao Y., Gao C., Wu H., Li Y., Cui Y., Guo X., Zheng S., Wang B., Yu K., Liang Q., Yang W., Lou X., Chen J., Feng M., Jian J., Zhang X., Luo G., Jiang Y., Liu J., Wang Z., Sha Y., Zhang B., Wu H., Tang D., Shen Q., Xue P., Zou S., Wang X., Liu X., Wang F., Yang Y., An X., Dong Z., Zhang K., Zhang X., Luo M. C., Dvorak J., Tong Y., Wang J., Yang H., Li Z., Wang D., Zhang A., WangJ. Draft genome of the wheat A-genome progenitor Triticum urartu. Nature. 2013; 496: 87-90.
- Jia J., Zhao S., Kong X., Li Y., Zhao G., He W., Appels R., Pfeifer M., Tao Y., Zhang X., Jing R., Zhang C., Ma Y., Gao L., Gao C., Spannagl M., Mayer K. F., Li D., Pan S., Zheng F., Hu Q., Xia X., Li J., Liang Q., Chen J., Wicker T., Gou C., Kuang H., He G., Luo Y., Keller B., Xia Q., Lu P., Wang J., Zou H., Zhang R., Xu J., Gao J., Middleton C., Quan Z., Liu G., Wang J., Yang H., Liu X., He Z., Mao L., Wang J. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature. 2013; 496: 91-95.
- Agarwal P., Baranwal V. K., Khurana P. Genome-wide Analysis of bZIP Transcription Factors in wheat and Functional Characterization of a TabZIP under Abiotic Stress.Rep. 2019; 9: 4608.
- Fowler S., Thomashow M. F. Arabidopsis transcriptome profiling indicates multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold-response pathway. Plant Cell. 2014; 14: 1675–1690.
- Chauhan H., Khurana N., Tyagi A. K., Khurana J. P., Khurana P. Identification and characterization of high-temperature stress-responsive genes in bread wheat (Triticum aestivum) and their regulation at various stages of development. Plant Mol.Biol. 2011; 5: 35–51.
- Kreps J. A., Wu Y., Chang H. S., Zhu T., Wang X., Harper J. F. Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress.Plant Physiol. 2002; 130: 2129–2141.
- Seki M., Ishida J., Narusaka M., Fujita M., Nanjo T., Umezawa T., Kamiya A., Nakajima M. Monitoring the expression pattern of around 7000 Arabidopsis genes under ABA treatments using a full-length cDNA microarray.Funct.Integr.Genom. 2002; 2: 282–291.
- Wong C. E., Li Y., Labbe A., Guevara D., Nuin P. Transcriptional profiling implicates novel interactions between abiotic stress and hormonal responses in Thellungiella, a close relative of Plant Physiol. 2006; 140: 1437–1450.
- Larkindale J., Hall J. D., Knight M. R., Vierling E. Heat stress phenotypes of Arabidopsis mutants implicate multiple signaling pathways in the acquisition of thermotolerance. Plant Physiol. 2005; 138: 882–897.
- Larkindale J., Vierling E. Core genome responses involved in acclimation to high temperature. Plant Physiol. 2008; 146(2): 748–761.
- Liu H.T., Li B., Shang Z. L., Li X. Z., Mu R. L., Sun D. Y., Zhou R. G. Calmodulin is involved in heat shock signal transduction in wheat. Plant Physiol. 2003; 132 (3): 1186–1195.
- Calderwood S. K., Xie Y., Wang X., Khaleque M. A., Chou S. D., Murshid A., Prince T., Zhang Y. Signal transduction always leading to heat-shock transcription. Signal Transduct. 2010; 2: 13-24
- Rampino P., Mita G., Fasano P., Borrelli G. M., Aprile A., Dalessandro G., De Bellis L., Perrotta C. Novel durum wheat genes upregulated in response to a combination of heat and drought stress. Plant Physiol.Biochem. 2012; 56: 72–78.
- Xin M., Wang Y., Yao Y., Xie C., Peng H., Ni Z., Sun Q. Diverse set of microRNAs are responsive to powdery mildew infection and heat stress in wheat (Triticum aestivum). BMC Plant Biol. 2010; 10: 123.
- Ragupathy R., Ravichandran S., Mahdi M. S., Huang D., Reimer E., Domaratzki M., Cloutier S. Deep sequencing of wheat sRNAtranscriptome reveals distinct temporal expression pattern of miRNAs in response to heat, light and UV. Rep. 2016; 6: 39373.
- Kumar R. R., Pathak H., Sharma S. K., Kala Y. K., Nirjal M. K., Singh G. P., Goswami S., Rai R. D. Novel and conserved heat-responsive microRNAs in wheat (Triticum aestivum). Funct.Integr.Genom.2015; 15: 323–348
- Rangan P., Furtado A., Henry., R. Transcriptome profiling of wheat genotypes under heat stress during grain-filling. Cereal Sci. 2020; 91: 102895.
- Nakamura S., Abe F., Kawahigashi H., Nakazono K., Tagiri A., Matsumoto T., Utsugi S., Ogawa T., Handa H., Ishida H., Mori M., Kawaura K., Ogihara Y., Miura H. A wheat homolog of MOTHER OF FT AND TFL1 acts in the regulation of germination. Plant Cell. 2011; 23(9): 3215–3229.
- Placido D. F., Campbell T., Folsom J., Cui X., Kruger G. R., Baenziger P. S., Walia H. Introgression of novel traits from a wild wheat relative improves drought adaptation in wheat. Plant Physiol. 2013; 16: 1806–1819.
- Liu J., Feng L., Li J., He Z. Genetic and epigenetic control of plant heat responses. Front.Plant Sci. 2015; 6: 267.
- Hakeem K. R., Chandna R., Ahmad P., Iqbal M., Ozturk M. Relevance of proteomic investigations in plant abiotic stress physiology. OMICS J.Integr.Biol. 2012; 16(11): 621-635.
- Majoul T., Bancel E., Triboï , Hamida J., Branlard G. Proteomic analysis of the effect of heat stress on hexaploid wheat grain: characterization of heat-responsive proteins from non-prolamins fraction. Proteomics. 2004; 4(2): 505–513.
- Skylas D. J., Cordwell S. J., Hains P. G., Larsen M. R., Bassel D. J., Walsh B. J., Blumenthal C., Rathmell W., Copeland L., Wrigley C.W. Heat shock of wheat during grain filling: proteins associated with heat- tolerance. Cereal Sci. 2002; 35: 175-188.
- Chauhan H., Khurana N., Nijhavan A., Khurana P., Khurana P. The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant Cell Environ. 2012; 35: 1912-1931.
- Wang X., Dinler B. S., Vignjevic M., Jacobsen S., Wollenweber B. Physiological and proteome studies of responses to heat stress during grain filling in contrasting wheat cultivars. Plant Sci. 2015; 230: 33-55.
- Laino, P., Shelton, D., Finnie, C., De Leonardis A.M., Mastrangelo A.M., Svensson B., Lafiandra D., Masci S. Comparative proteome analysis of metabolic proteins from seeds of durum wheat (cv. Svevo) subjected to heat stress. J Proteomics. 2010; 10: 2359–2368.
- Yang F., Jørgensen A. D., Li H., Søndergaard I., Finnie C., Svensson B., Jiang D., Wollenweber B., Jacobsen S. Implications of high-temperature events and water deficits on protein profiles in wheat (Triticum aestivum cv. Vinjett) grain. Proteomics. 2011; 11: 684–1695.
- Xue G. P., Drenth J., McIntyre C. L. TaHsfA6f is a transcriptional activator that regulates a suite of heat stress protection genes in wheat (Triticum aestivum) including previously unknown Hsf targets. J. Exp. Bot. 2015; 66: 1025–1039.
- Lu Y., Li R., Wang R., Wang X., Zheng W., Sun Q., Tong S., Dai S., Xu S. Comparative proteomic analysis of flag leaves reveals new insight into wheat heat adaptation.Plant Sci. 2017; 8: 1–11.
- Kumar R. R., Singh G. P., Goswami S., Pathak H., Rai R. D. Proteome analysis of wheat (Triticum aestivum) for the identification of differentially expressed heat-responsive proteins. J.Crop Sci. 2014; 8: 973–986.
- Gupta O. P., Sharma I. Deciphering the dynamics of changing proteins of tolerant and intolerant wheat seedlings subjected to heat stress. Biol.Rep. 2015; 42: 43–51.
- Kaplan F., Kopka J., Haskell D.W., Zhao W., Cameron Schiller K., Gatzke N., Sung D.Y., Guy C. L. Exploring the temperature-stress metabolome of Arabidopsis. Plant Physiol. 2004; 136: 4159–4168.
- Schauer N., Fernie A. R. Plant metabolomics: towards biological function and mechanism. Trends Plant Sci. 2006; 11: 508–516.
- Putri S. P., Yamamoto S., Tsugawa H., Fukusaki E. Current metabolomics: technological advances. Biosci. Bioeng. 2013; 116: 9–16.
- Fernie A. R., Aharoni A., Willmitzer L., Stitt M., Tohge T., Kopka J., Carroll A. J., Saito K., Fraser P. D., DeLuca V. Recommendations for reporting metabolite data. Plant Cell. 2011; 23: 2477–2482.
- de Leonardis A. M., Fragasso M., Beleggia R., Ficco D. B., de Vita P., Mastrangelo A. M. Effects of heat stress on metabolite accumulation and composition, and nutritional properties of durum wheat grain. J.Mol.Sci. 2015; 16(12): 30382–30404
- Grover A., Mittal D., Negi M., Lavania D. Generating high-temperature tolerance transgenic plants: achievements and challenges. Plant Sci. 2013; 205: 38–47.
- Pellegrineschi A, Reynolds M, Pacheco M, Brito R. M, Almeraya R, Yamaguchi-Shinozaki K, Hoisington D. Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome. 2004; 47: 493–500.
- Rong W., Qi L., Wang A., Ye X., Du L., Liang H., Xin Z., Zhang Z. The ERF transcription factor TaERF3 promotes tolerance to salt and drought stresses in wheat. Plant Biotechnol.J. 2014; 12: 468–479.
- Saad A. S., Li X., Li H. P., Huang T., Gao C. S., Guo M. W., Cheng W., Zhao G.Y., Liao Y.C. A rice stress-responsive NAC gene enhances tolerance of transgenic wheat to drought and salt stresses. Plant Sci. 2013; 203–204: 33–40.
- Lata C., Prasad M. Role of DREBs in regulation of abiotic stress responses in plants. Exp. Bot. 2011; 62: 4731-4748.
- Fu J., Momčilović I., Clemente T., Nersesian N., Trick H. N., Ristic Z. Heterologous expression of a plastid EF-Tu reduces protein thermal aggregation and enhances CO2 fixation in wheat (Triticum aestivum) following exposure to heat stress. Plant Mol.Biol. 2008; 68: 277–288.
- Guo W., Zhang J., Zhang N., Xin M., Peng H., Hu Z., Ni Z., Du J. The wheat NAC transcription factor TaNAC2L is regulated at the transcriptional and post-translational levels and promotes heat stress tolerance in transgenic PLoS One.2008; 10(8) e0135667.
- He G. H., Xu J. Y., Wang Y. X., Liu J. M., Li P. S., Ming C., Ma Y. Z., Xu Z. S. Drought-responsive WRKY transcription factor genes TaWRKY1 and TaWRKY33 from wheat confer drought and/or heat resistance in Arabidopsis. BMC Plant Biol. 2016; 16: 116.
- El-Esawi M. A., Al-Ghamdi A. A., Ali H. M., Ahmad M. Overexpression of AtWRKY30 Transcription Factor Enhances Heat and Drought Stress Tolerance in Wheat (Triticum aestivum). Genes (Basel). 2019; 10(2):163.
- Queitsch C., Hong S. W., Vierling E., Lindquest S. Heat shock protein 101 plays a crucial role in thermotolerance in Arabidopsis. Plant Cell. 2000; 12: 479–492.
- Hong S.W., Vierling E. Mutants of Arabidopsis thaliana defective in the acquisition of tolerance to high temperature stress. Natl. Acad. Sci. 2000; 97: 4392–4397.
- Nieto-Sotelo J., Martınez L. M., Ponce G. Maize HSP101 plays important roles in both induced and basal thermo tolerance and primary root growth. Plant Cell. 2002; 14: 1621–1633.
- Wu C. Heat shock transcription factors: structure and regulation. Rev. Cell Dev. Biol. 1995; 11: 441–469.
- Agarwal M., Sarkar N., Grover A. Low molecular weight heat shock proteins in plants. Journal of Plant Biology. 2003; 30: 141–149.
- Kumar R. R., Goswami S., Sharma S. K. Differential expression of heat shock protein and alteration in osmolyte accumulation under heat stress in wheat. Plant Biochem.Biotechnol. 2013; 22: 16–26.
- Chauhan H., Khurana N., Nijhavan A., Khurana J. P., Khurana P. The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant Cell Environ. 2012; 35: 1912-1931.
- Nieto-Sotelo J., Martınez L. M., Ponce G. Maize HSP101 plays important roles in both induced and basal ther motolerance and primary root growth. Plant Cell. 2002; 14: 1621–1633.
- Malik M. K., Slovin J. P., Hwang C. H., Zimmerman J. L. Modified expression of a carrot small heat shock protein gene, hsp17.7, results in increased or decreased thermotolerance. Plant J. 1999; 20: 89–99.
- Chauhan H., Khurana N., Agarwal P., Khurana J. P., Khurana P. A Seed Preferential Heat Shock Transcription Factor from Wheat Provides Abiotic Stress Tolerance and Yield Enhancement in Transgenic Arabidopsisunder Heat Stress Environment. PLoS ONE. 2013; 8(11) e79577.
- Hu, X.J., Chen, D., Lynne, M.C., Fernanda, D.M., Zhang, Z.B., Drenth, J., Kalaipandian, S., Chang, H., Xue, G.P. Heat shock factor C2a serves as a proactive mechanism for heat protection in developing grains in wheat via an ABA-mediated regulatory pathway: TaHsfC2a plays a proactive heat protection role. Plant Cell Environ. 2018; 41: 79–98.
- Farooq M., Wahid A., Kobayashi N., Fujita D., Basra S. M. A. Plant drought stress: effects, mechanisms and management. Agron Sustain Dev. 2009; 29: 185–212.
- Yang X., Liang Z., Lu C. Genetic engineering of the biosynthesis of glycine betaine enhances photosynthesis against high-temperature stress in transgenic tobacco plants. Plant Physiol. 2005; 138: 2299–2309.
- Wang G., Hui Z., Li F. Improvement of heat and drought photosynthetic tolerance in wheat by overaccumulation of glycine betaine. Plant Biotechnol.Rep. 2010; 4: 213–222.
- Suryavanshi P., Buttar G. S. Mitigating terminal heat stress in wheat. j. bio-resour. stress manag. 2016; 7(1): 142-150.
- Shi W. M., Muramoto Y., Ueda A., Takabe T. Cloning of peroxisomal ascorbate peroxidase gene from barley and enhanced thermotolerance by overexpressing in Arabidopsis thaliana. Gene. 2000; 273: 23–27.
- Tang L., Kwon S.Y., Kim S.H. Enhanced tolerance of transgenic potato plants expressing both superoxide dismutase and ascorbate peroxidase in chloroplasts against oxidative stress and high temperature. Plant Cell Rep. 2006; 25: 1380–1386.
- Chen J., Wang P., Mi H. L., Chen G.Y., Xu D. Q. Reversible association of ribulose-1, 5-bisphosphate carboxylase/oxygenase activase with the thylakoid membrane depends upon the ATP level and pH in rice without heat stress. Exp. Bot. 2010; 61: 2939–2950.
- Murakami Y., Tsuyama M., Kobayashi Y., Kodama H., Iba K. Trienoic fatty acids and plant tolerance of high temperature. Science. 2000; 287: 476–479.
- Wang F., Zang X. S., Kabir M. R., Liu K. L., Liu Z. S., Ni Z. F., Yao Y. Y., Hu Z. R., Sun Q. X., Peng H. R. A wheat lipid transfer protein 3 could enhance the basal thermo tolerance and oxidative stress resistance of Arabidopsis. Gene. 2014; 550: 18–26.
- Singh A., Khurana P. Molecular and functional characterization of a wheat B2 protein imparting adverse temperature tolerance and influencing plant growth. Plant Sci. 2016; 7: 642.
- Zhang L., Geng X., Zhang H., Zhou C., Zhao A., Wang F., Zhao Y., Tian X., Hu Z., Xin M. Isolation and characterization of heat-responsive gene TaGASR1 from wheat (Triticum aestivum). J.Plant Biol. 2017; 60: 57–65.
- Zang X., Geng X., Liu K., Fei W., Liu Z., Zhang L., Yue Z., Tian X., Hu Z., Yao Y. Ectopic expression of TaOEP16-2-5B, a wheat plastid outer envelope protein gene, enhances heat and drought stress tolerance in transgenic Arabidopsis Plant Science. 2017a; 258: 1–11.
- Zang X., Geng X., Wang F., Liu Z., Zhang L., Zhao Y., Tian X., Ni Z., Yao Y., Xin M. Overexpression of wheat ferritin gene TaFER-5B enhances tolerance to heat stress and other abiotic stresses associated with the ROS scavenging. BMC Plant Biol. 2017b; 17: 14.
- Zang X., Geng X., He K., Wang F., Tian X., Xin M., Yao Y., Hu Z., Ni Z., Sun Q., Peng H. Overexpression of the Wheat (Triticum aestivum ) TaPEPKR2 Gene Enhances Heat and Dehydration Tolerance in Both Wheat and Arabidopsis. Front.Plant Sci. 2018; 9: 1710.
- Liu Z., Xin M., Qin J., Peng H Ni Z., Yao Y., Sun Q. Temporal transcriptome profiling reveals expression partitioning of homoeologous genes contributing to heat and drought acclimation in wheat (Triticum aestivum ). BMCPlant Biol. 2015; 15: 1–20.
- Gardiner L. J., Quintontulloch M., Olohan L., Price J., Hall N., Hall A. A genome-wide survey of DNA methylation in hexaploid wheat. Genome Biol. 2015; 16: 273.
- Ni Z., Li H., Zhao Y., Peng H., Hu Z., Xin M., Sun Q. Genetic improvement of heat tolerance in wheat: Recent progress in understanding the underlying molecular mechanisms. Crop J. 2018; 6: 32–41.
- Charon C., Moreno A. B., Bardou F., Crespi M. Non-protein-coding RNAs their interacting RNA-binding proteins in the plant cell nucleus. Mol Plant. 2010; 3: 729–739.
- Xin M., Wang Y., Yao Y., Xie C., Peng H., Ni Z., Sun Q. Diverse set of microRNAs are responsive to powdery mildew infection and heat stress in wheat (Triticum aestivum). BMC Plant Biol. 2010; 10: 123.\
- Giusti L., Mica E., Bertolini E., De Leonardis A. M., Faccioli P., Cattivelli L., Crosatti C. microRNAs differentially modulated in response to heat and drought stress in durum wheat cultivars with contrasting water use efficiency. Integr.Genom. 2017; 17(2-3) 293–309.
- Wang Y., Sun F., Cao H., Peng H Ni Z., Sun Q., Yao Y. TamiR159 directed wheat TaGAMYB cleavage and its involvement in anther development and heat response. PLoS One. 2012;7(11) :e48445.
- Ravichandran S., Ragupathy R., Edwards T., Domaratzki M, Cloutier S. MicroRNA-guided regulation of heat stress response in wheat. BMC Genomics. 2019; 20: 488.
- Acuñagalindo M. A., Mason R. E., Subramanian N. K., Hays D. B. Meta-analysis of wheat QTL regions associated with adaptation to drought and heat stress. Crop Sci. 2015; 55: 477–492.
- Mondal S., Mason R. E., Huggins T., Hays D. B. QTL on wheat (Triticum aestivum) chromosomes 1B, 3D and 5A are associated with constitutive production of leaf cuticular wax and may contribute to lower leaf temperatures under heat stress. Euphytica. 2015; 201 : 123–130.